
Publications
You can find a full and up-to-date list of our publications on PubMed. Hover over each image for a short summary of each publication.
-
 Mäeots et al. report the development of the Chronobot; an AI-integrated time-resolved cryo-EM plunge freezing machine.
Mäeots et al. report the development of the Chronobot; an AI-integrated time-resolved cryo-EM plunge freezing machine.Mäeots ME, Tupin S, Esfahani NMH, Rodriguez-Molina JB, Clapperton JA, Amin A, Imbert A, Enchev RI.
Chronobot: Deep learning guided time-resolved cryo-EM captures molecular choreography of RecA in homology search
BioRXiv (2025). DOI -
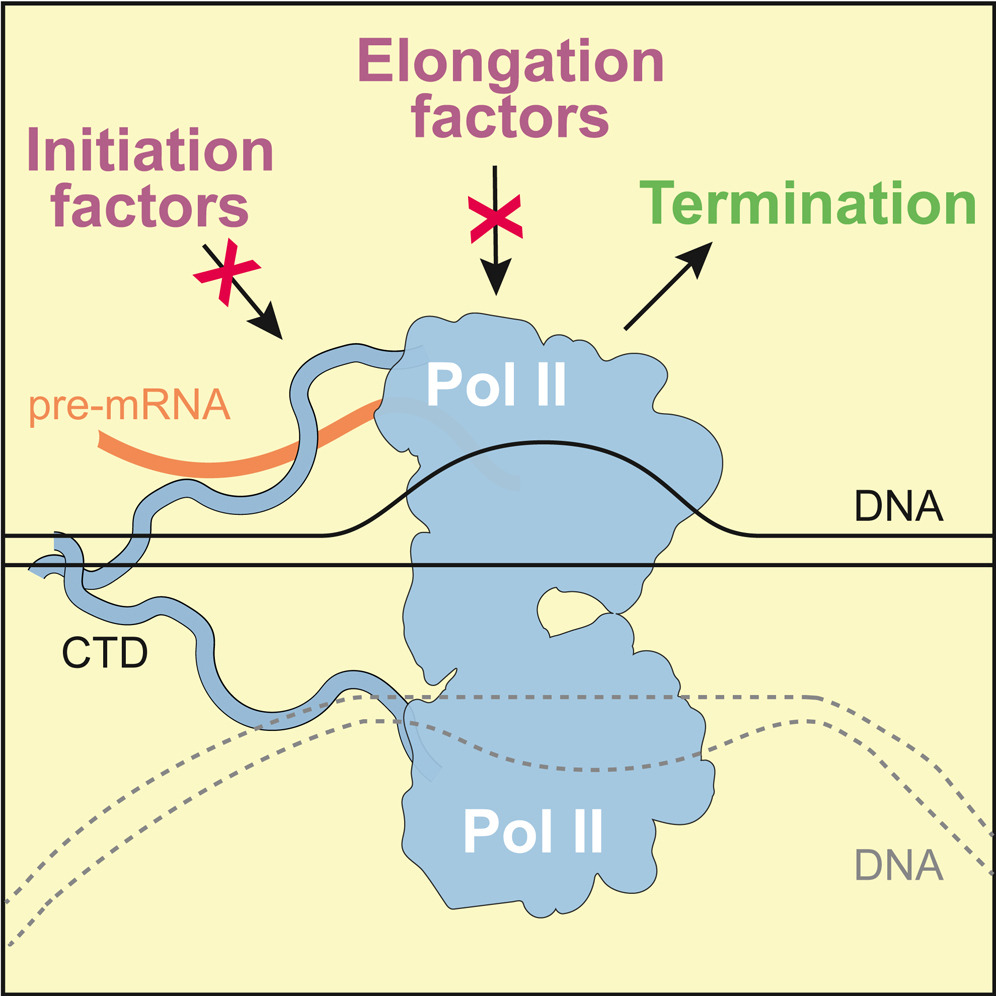 Carminati et al. report a direct interaction between the 3'-end processing machinery and Pol II, and a regulated dimerzation of Pol II that may be important in transcription termination.
Carminati et al. report a direct interaction between the 3'-end processing machinery and Pol II, and a regulated dimerzation of Pol II that may be important in transcription termination.Carminati M, Rodríguez-Molina JB, Manav MC, Bellini D, Passmore LA.
A direct interaction between CPF and RNA Pol II links RNA 3' end processing to transcription
Molecular Cell (2023). DOI -
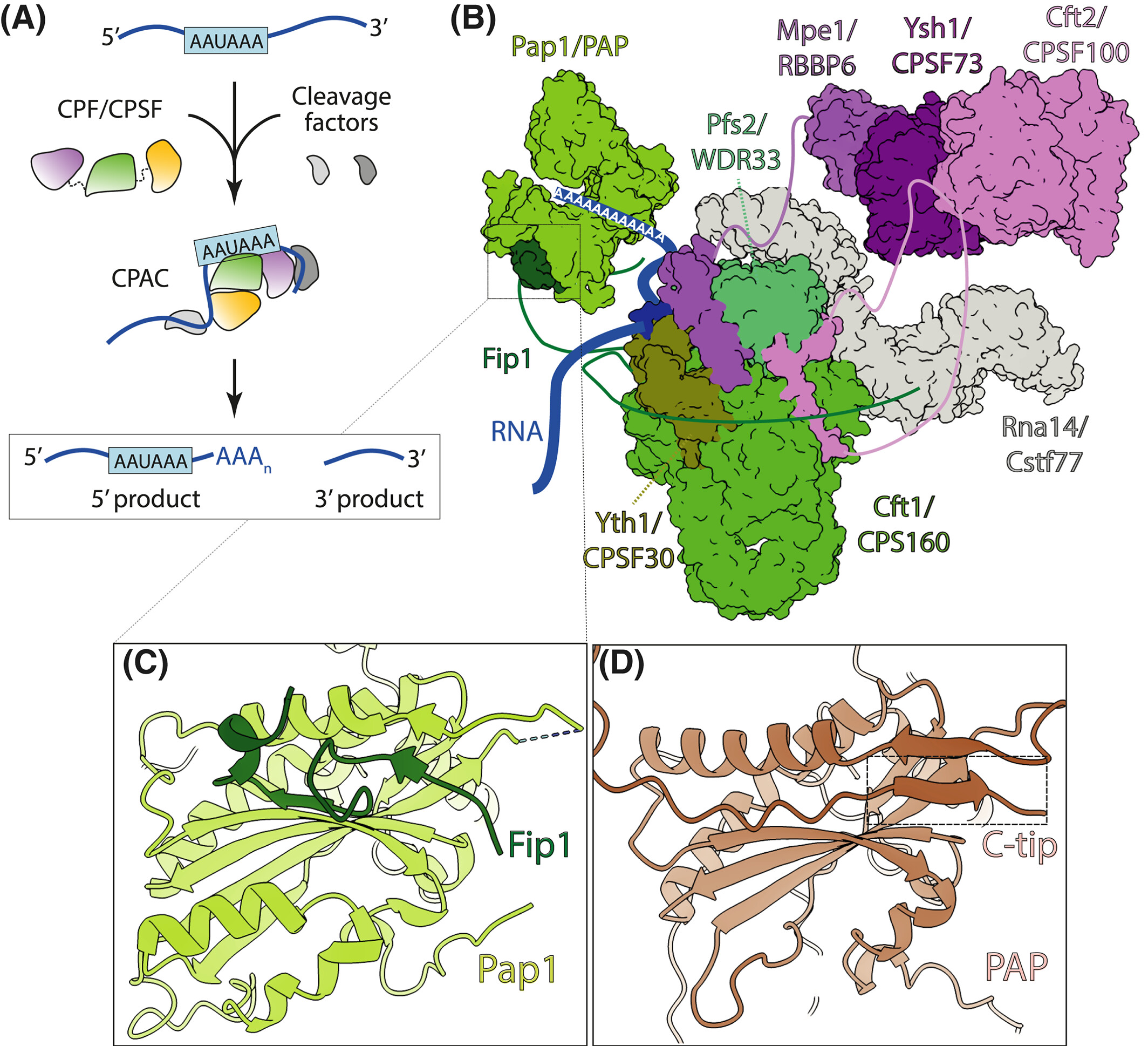 Here we review the literature on regulatory mechanisms of mRNA polyadenylation.
Here we review the literature on regulatory mechanisms of mRNA polyadenylation.Rodríguez-Molina JB, Turtola M.
Birth of a poly(A) tail: mechanisms and control of mRNA polyadenylation
FEBS Open Bio. (2023). DOI -
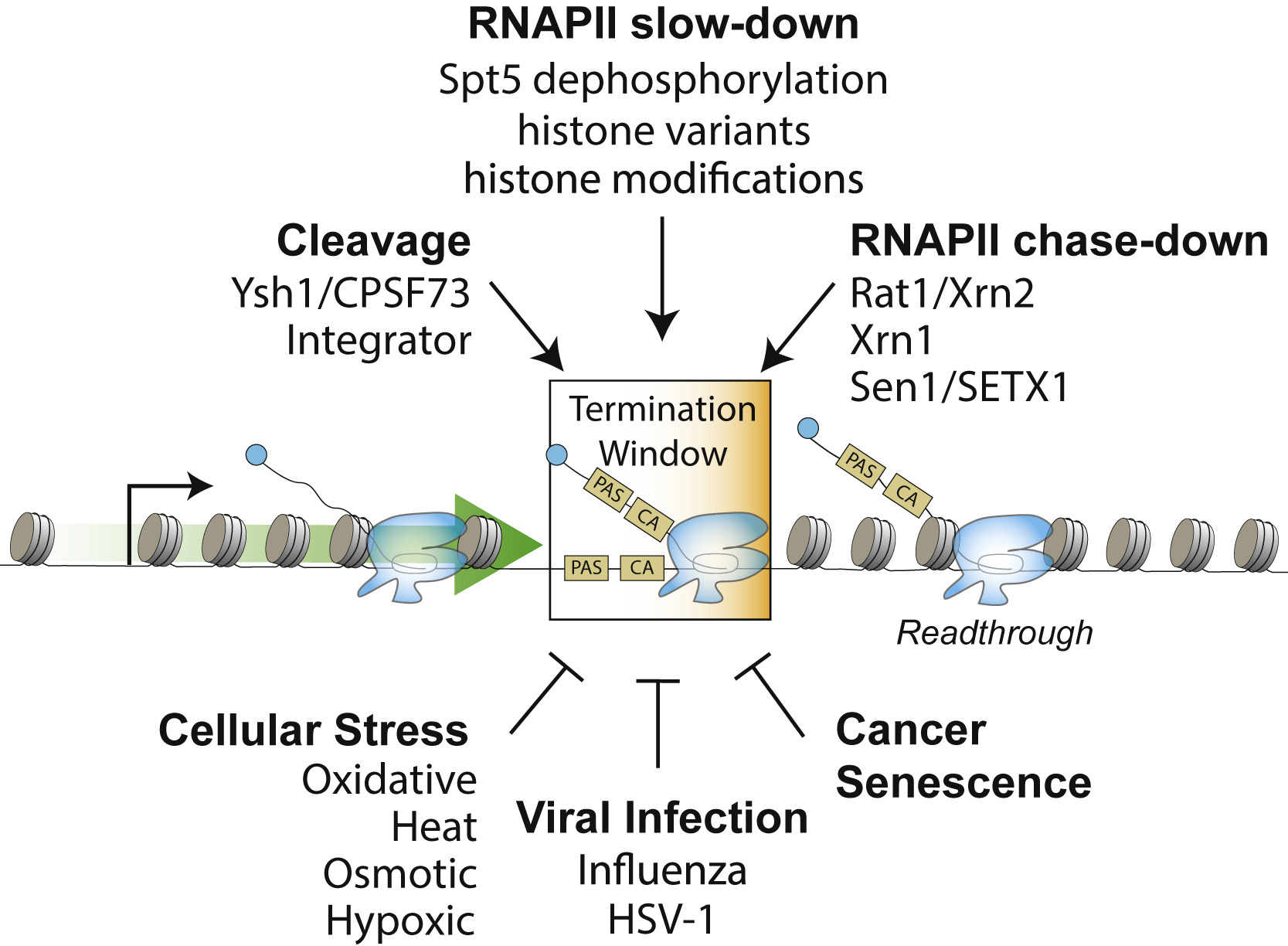 Here we review the literature on models of Pol II transcription termination, its coupling to RNA processing, and its regulation in various contexts.
Here we review the literature on models of Pol II transcription termination, its coupling to RNA processing, and its regulation in various contexts.Rodríguez-Molina JB, West S, Passmore LA.
Knowing when to stop: Transcription termination on protein-coding genes by eukaryotic RNAPII
Molecular Cell (2023). DOI -
 We report that Mpe1 functions as an RNA binding sensor to activate CPF cleavage activity, regulate polyadenyltion, and terminate transcription.
We report that Mpe1 functions as an RNA binding sensor to activate CPF cleavage activity, regulate polyadenyltion, and terminate transcription.Rodríguez-Molina JB, O'Reilly FJ, Fagarasan H, Sheekey E, Maslen S, Skehel JM, Rappsilber J, Passmore LA.
Mpe1 senses the binding of pre-mRNA and controls 3' end processing by CPF
Molecular Cell (2022). DOI -
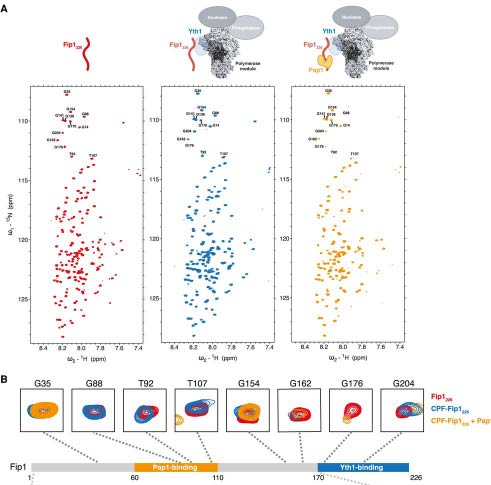 Kumar et al. use biochemistry and NMR to show that the flexibility of the Fip1 subunit of CPF is important for cleavage and polyadenylation of pre-mRNAs.
Kumar et al. use biochemistry and NMR to show that the flexibility of the Fip1 subunit of CPF is important for cleavage and polyadenylation of pre-mRNAs.Kumar A, Yu CWH, Rodríguez-Molina JB, Li XH, Freund SMV, Passmore LA.
Dynamics in Fip1 regulate eukaryotic mRNA 3' end processing
Genes & Development (2021). DOI -
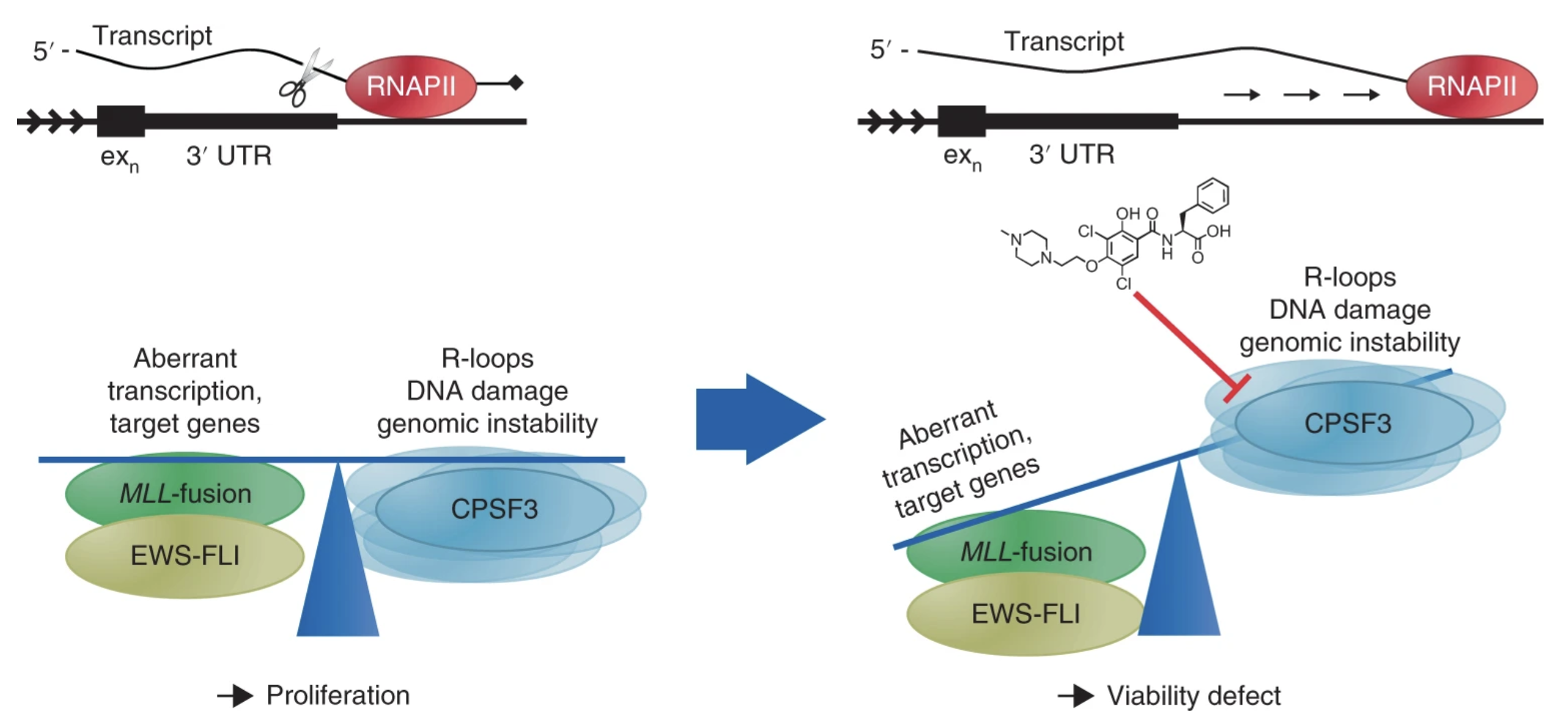 Ross et al. characterise JTE-607 as an inhibitor of CPSF73 activity with anti-cancer properties.
Ross et al. characterise JTE-607 as an inhibitor of CPSF73 activity with anti-cancer properties.Ross NT, Lohmann F, Carbonneau S, Fazal A, Weihofen WA, Gleim S, Salcius M, Sigoillot F, Henault M, Carl SH, Rodríguez-Molina JB, et al.
CPSF3-dependent pre-mRNA processing as a druggable node in AML and Ewing's sarcoma
Nature Chemical Biology (2020). DOI -
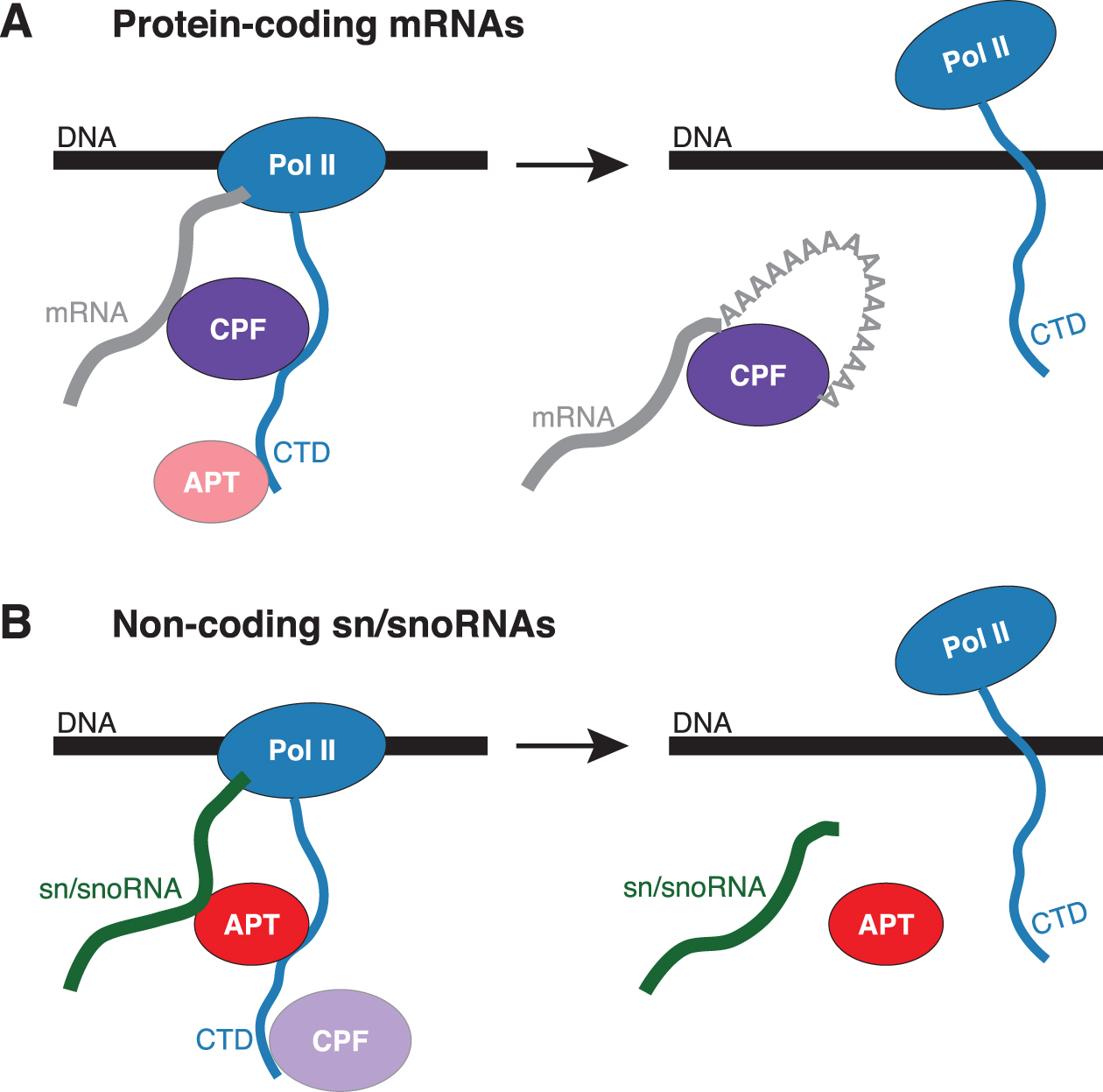 Lidschreiber et al. report CPF and APT are distinct complexes in yeast that function in processing of mRNAs and sn/snoRNAs, respectively.
Lidschreiber et al. report CPF and APT are distinct complexes in yeast that function in processing of mRNAs and sn/snoRNAs, respectively.Lidschreiber M, Easter AD, Battaglia S, Rodríguez-Molina JB, Casañal A, Carminati M, Baejen C, Grzechnik P, Maier KC, Cramer P, Passmore LA.
The APT complex is involved in non-coding RNA transcription and is distinct from CPF
Nucleic Acids Research (2018). DOI -
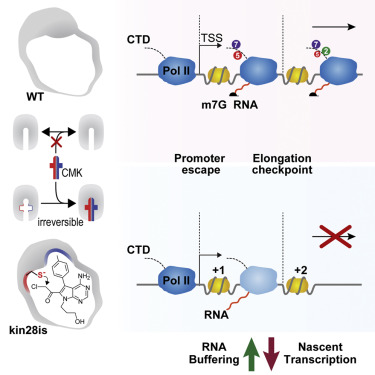 We report a new strategy for the covalent chemical inhibition of protein kinases in vivo. We show that the Pol II CTD kinase, Kin28, is important for Pol II to overcome an elongation checkpoint near the +2 nucleosome.
We report a new strategy for the covalent chemical inhibition of protein kinases in vivo. We show that the Pol II CTD kinase, Kin28, is important for Pol II to overcome an elongation checkpoint near the +2 nucleosome.Rodríguez-Molina JB, Tseng SC, Simonett SP, Taunton J, Ansari AZ.
Engineered Covalent Inactivation of TFIIH-Kinase Reveals an Elongation Checkpoint and Results in Widespread mRNA Stabilization
Molecular Cell (2016). DOI -
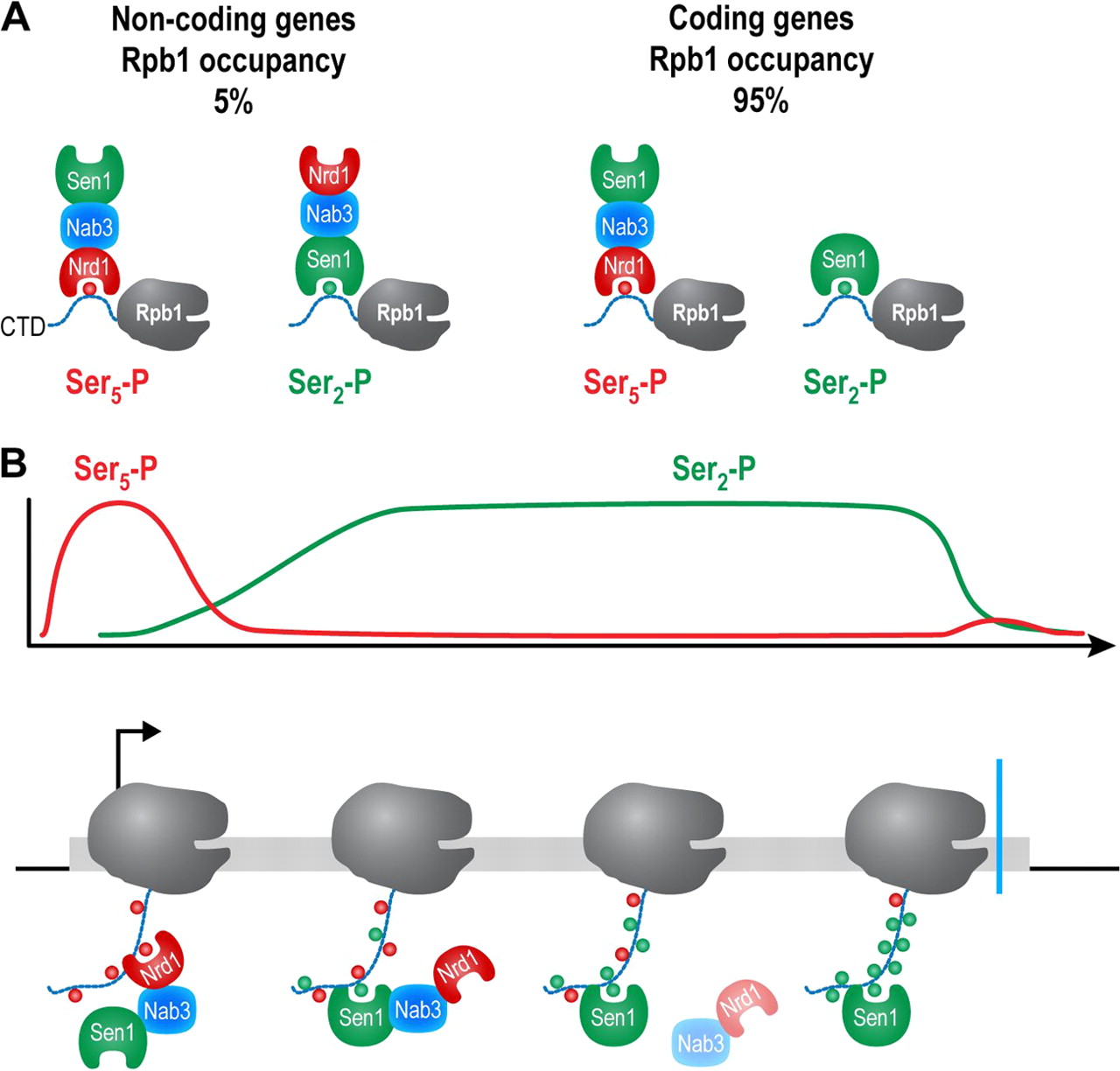 Chinchilla et al. report an interaction between the Sen1 helicase and the Ser2-phosphorylated CTD of Pol II. This interaction may be important for Sen1 recruitment at specific gene classes.
Chinchilla et al. report an interaction between the Sen1 helicase and the Ser2-phosphorylated CTD of Pol II. This interaction may be important for Sen1 recruitment at specific gene classes.Chinchilla K, Rodriguez-Molina JB, Ursic D, Finkel JS, Ansari AZ, Culbertson MR.
Interactions of Sen1, Nrd1, and Nab3 with multiple phosphorylated forms of the Rpb1 C-terminal domain in Saccharomyces cerevisiae
Eukaryotic Cell (2012). DOI -
 Zhang et al. report Ssu72 as a Ser7 Pol II CTD phosphatase and its role in global trancription regulation.
Zhang et al. report Ssu72 as a Ser7 Pol II CTD phosphatase and its role in global trancription regulation.Zhang DW, Mosley AL, Ramisetty SR, Rodríguez-Molina JB, Washburn MP, Ansari AZ.
Ssu72 phosphatase-dependent erasure of phospho-Ser7 marks on the RNA polymerase II C-terminal domain is essential for viability and transcription termination
Journal of Biological Chemistry (2012). DOI -
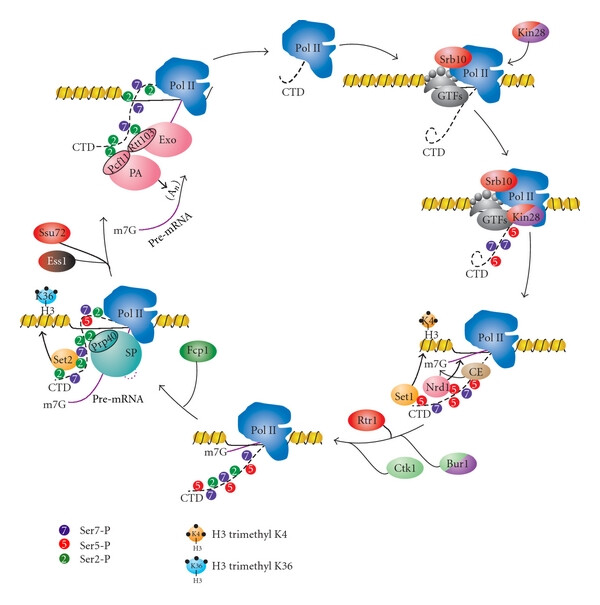 Here we review the role of the Pol II CTD in coordinating transcription with RNA processing and chromatin modifications.
Here we review the role of the Pol II CTD in coordinating transcription with RNA processing and chromatin modifications.Zhang DW, Rodríguez-Molina JB, Tietjen JR, Nemec CM, Ansari AZ.
Emerging Views on the CTD Code
Genetics Research International (2012). DOI -
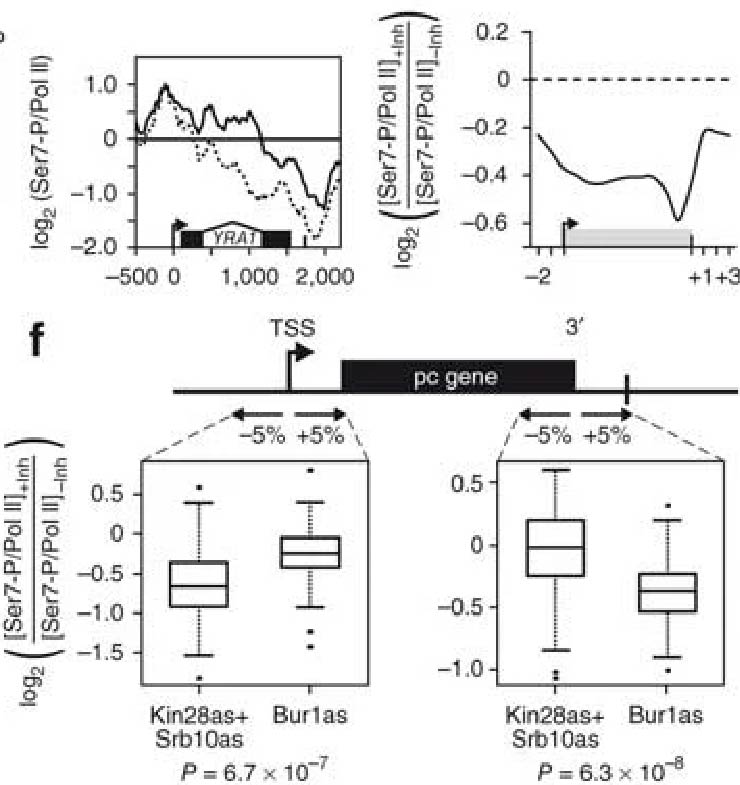 Tietjen et al. report widespread occupancy of phospho-Ser7 Pol II CTD marks and the role of Bur1/CDK9 in placing this mark promoter-distally.
Tietjen et al. report widespread occupancy of phospho-Ser7 Pol II CTD marks and the role of Bur1/CDK9 in placing this mark promoter-distally.Tietjen JR, Zhang DW, Rodríguez-Molina JB, White BE, Akhtar MS, Heidemann M, Li X, Chapman RD, Shokat K, Keles S, Eick D, Ansari AZ.
Chemical-genomic dissection of the CTD code
Nature Structure & Molecular Biology (2010). DOI